XI
L’ORGANIZZAZIONE FISIOLOGICA DELLE EMICRANIE
Le ventiquattro lettere dell’alfabeto formano, nelle diverse lingue, una varietà di parole non superiore alla gamma dei sintomi prodotti nelle diverse persone dai pensieri indotti dalla melanconia. Essi sono irregolari, oscuri, vari, così infiniti, da superare lo stesso Proteo per varietà d’aspetti ...
ROBERT BURTON
Come Burton aveva osservato nel caso della melanconia, anche per l’emicrania il problema sta nell’infinita varietà di sintomi che siamo chiamati a spiegare. Dobbiamo tentare di formulare una teoria, o un insieme di teorie, che siano nel complesso più generali dei semplici meccanismi visti nel precedente capitolo: abbastanza generali da coprire ogni aspetto di qualunque tipo di emicrania, e tuttavia applicabili in modo specifico a ogni sintomo particolare.
Cominceremo considerando i sintomi dell’emicrania a diversi livelli funzionali, dal più basso al più alto, e i processi che possono trovarsi alla base di tali sintomi. Se questi metodi si riveleranno inadeguati, saremo costretti ad adottare concetti radicalmente diversi su cosa si intenda per «funzione» o «centro» in campo neurofisiologico, e a pensare in termini di organizzazione dinamica di sistemi funzionali, piuttosto che nei termini tradizionali di apparato neurale e di processi fissi.
Si è visto che i sintomi vegetativi dell’emicrania esprimono una predominanza parasimpatica e non possiamo fare a meno di restar colpiti dal fatto che la diversa importanza dei diversi sintomi sia abbinata alla configurazione anatomica e funzionale del sistema nervoso parasimpatico. Dal punto di vista anatomico, esistono plessi gangliari ben distinti in tutte le principali strutture viscerali, vascolari e ghiandolari dell’organismo, ed essi costituiscono il livello più basso di rappresentazione neurale dei visceri. Possiamo ipotizzare che l’esistenza di molti di questi plessi funzionalmente discreti fornisca una base fisiologica per molte sindromi parasimpatiche distinte, e che l’attività neuronica locale in tali plessi, una volta scatenata da una scarica iniziale proveniente dal sistema nervoso centrale, possa persistere (per minuti, ore, o giorni) come un’eccitazione funzionalmente discreta e separata. Nel caso di un paziente con un’emicrania protratta e localizzata, ad esempio, possiamo immaginare un disturbo relativamente isolato confinato in un singolo plesso perivascolare; in altri casi, l’eccitazione neuronica locale può essere localizzata nei plessi intramurali dello stomaco, del colon, delle ghiandole lacrimali, ecc. Questa prima alterazione neuronica può in seguito essere rinforzata da modificazioni tessutali locali; per esempio, dalla produzione di essudato e dall’infiammazione sterile che si osserva nelle cefalee vascolari protratte, o da altre modificazioni a lungo termine che fanno seguito al disturbo neurogeno iniziale. I sintomi vegetativi dell’emicrania, al livello più basso, possono essere mediati, classificati e isolati grazie alla configurazione a rete nervosa del sistema parasimpatico. Le «istruzioni» sulla forma dell’attacco, o sul tipo di equivalente emicranico, sono presumibilmente determinate da meccanismi centrali, sebbene possano anche intervenire, in modo accessorio, differenze di soglia locali, innate o condizionate. È tipico, comunque, che quando un organo bersaglio, o un plesso locale, viene rimosso dal sistema (ad esempio, per effetto di un intervento chirurgico), gli attacchi di emicrania si ripresentino, in una forma leggermente alterata. Questo suggerisce che i meccanismi periferici prefissati vengano usati in base alla loro disponibilità, grazie al fatto che l’organizzazione centrale dell’emicrania è plastica e flessibile.
Il termine «parasimpatico» fu originariamente usato, e viene opportunamente limitato, a indicare strutture e attività periferiche. È chiaro che non basta parlare dell’emicrania come di un attacco parasimpatico, in quanto tutti gli attacchi hanno anche componenti centrali, cerebrali. Utili e appropriati si sono rivelati i concetti e i termini ideati da Hess, nei suoi famosi studi sulla funzione vegetativa e diencefalica (Hess, 1954). Hess chiama «ergotropica» la combinazione di un’attività simpatica periferica con un risveglio centrale, e «trofotropica» la condizione inversa. Questi termini non sono solo fisiologici, ma anche biologici, nel senso che si riferiscono all’organismo nel suo complesso. Così, l’ergotropia indica la tendenza di un organismo a rivolgersi verso il mondo esterno, a essere operativo, a compiere del lavoro: tutte attività per le quali esso sarà dotato di un’accresciuta vigilanza e acuità sensoria, di un tono muscolare e di un tono simpatico potenziati. La trofotropia indica la tendenza di un organismo a rivolgersi verso l’interno, alla propria economia; perciò esso sarà dotato di attività viscerali e ghiandolari potenziate, associate all’inibizione del livello della coscienza, dell’acuità sensoria e del tono muscolare. La terminologia di Hess è straordinariamente utile per la nostra comprensione degli attacchi emicranici. È evidente che, nel corso della parte principale dell’attacco, sono rispettati tutti i suoi criteri di trofotropia (aumento del tono parasimpatico, affievolimento dell’allerta, ipersincronia dell’elettroencefalogramma, ecc.): possiamo dire che la parte principale di un’emicrania rappresenta una sindrome trofotropica polimorfa.
L’elaborazione sperimentale dei concetti di Hess, con l’applicazione a problemi clinici, è stata intrapresa da molti ricercatori, tra i quali spicca Ernst Gellhorn. È stata ad esempio dimostrata (si veda Gellhorn, 1967) l’esistenza, a tutti i livelli nel nevrasse, di sistemi ergotropici e trofotropici che sono anatomicamente, fisiologicamente e farmacologicamente distinti. È stato dimostrato che le attività ergotrope e trofotrope vengono mantenute normalmente in equilibrio reciproco; tale equilibrio determina quella che Gellhorn ha chiamato «sintonia» del sistema nervoso in ogni dato momento. Ad esempio, l’inibizione del sistema ergotropico è associata all’eccitazione della componente trofotropica, e viceversa. È tipico, inoltre, che qualsiasi marcato cambiamento di sintonia del sistema autonomo sarà seguito da un fenomeno di rimbalzo nella direzione opposta.
Possiamo ora tradurre la sequenza di un’emicrania comune nella terminologia di Hess. I sintomi prodromici o iniziali sono quelli della predominanza ergotropica; la parte principale dell’attacco rappresenta un collasso nella trofotropia; i sintomi del rimbalzo sono ancora ergotropici. Così, un’emicrania comune può essere considerata come un parossismo a tre stadi, in lenta evoluzione, durante il quale si verificano caratteristiche e coerenti modificazioni degli equilibri nervosi. Questo concetto corrisponde approssimativamente alla descrizione di Lennox dell’emicrania come «accesso vegetativo». Non possiamo stabilire a quale preciso livello il processo emicranico abbia origine, e in effetti sospettiamo che, posta in questi termini, la domanda sia priva di significato, poiché i sistemi ergotropico e trofotropico sono rappresentati gerarchicamente, attraverso tutto il nucleo del nevrasse, dalle corna spinali (intermedio-laterali) alla formazione reticolare del tronco encefalico, all’ipotalamo, fino a raggiungere le porzioni medio-basali della corteccia cerebrale.
Possiamo anche riconoscere un ciclo di eccitazione e inibizione nel decorso molto concentrato dell’aura emicranica, là dove il ciclo di eccitazione (scotomi scintillanti, parestesie, eccitamento, risveglio sensoriale diffuso, ecc.), quello di inibizione (scotomi negativi, anestesia, sonnolenza, mancamenti, sincope, inibizione sensoria diffusa e simili) e, eventualmente, il successivo ritorno all’eccitamento, richiedono solo da 30 a 40 minuti.
Arriviamo quindi a tracciare un quadro del processo emicranico che è simile a quello proposto da Liveing un secolo fa: una forma di accesso centroencefalico, la cui attività è proiettata rostralmente sugli emisferi cerebrali e perifericamente attraverso le ramificazioni del sistema nervoso autonomo. Possiamo immaginare la corteccia, nel corso di un’aura emicranica, sottoposta a un bombardamento ascendente, al quale essa risponde con proprie attività secondarie: queste sono multifocali (scotomi scintillanti, parestesie, ecc.) e si mettono in moto su uno sfondo caratterizzato da un risveglio corticale diffuso. In modo simile, possiamo riconoscere che i plessi autonomi periferici sono soggetti a un fuoco di fila discendente, al quale essi rispondono con attività multifocali secondarie indipendenti. Questa descrizione del processo emicranico è schematizzata nella figura 17.

Fig. 17. Schema dell’ipotetico processo emicranico. Questo è rappresentato come un’attività parossistica lenta, ciclica, centroencefalica, proiettata rostralmente verso la corteccia cerebrale, dove innesca i processi secondari dell’aura emicranica (scotomi, parestesie, ecc.) e caudalmente verso le ramificazioni dei plessi vegetativi attraverso l’intero organismo. Il processo è, secondo le parole di Gowers, «... molto misterioso ... vi è una forma peculiare di attività che sembra diffondersi come le increspature sulla superficie di uno specchio d’acqua nel quale sia stato gettato un sasso ... là dove sono passate le onde, rimane uno stato simile a un disturbo molecolare delle strutture».
È ovvio, tuttavia, che occorreranno altri concetti e altri termini per descrivere l’attivazione della corteccia cerebrale nel corso di un’aura emicranica. Le allucinazioni visive dell’emicrania ci forniscono le più chiare indicazioni di questi processi superiori e di come sono organizzati.
Si è già osservato (si veda il capitolo III) come vi sia di solito una sequenza di allucinazioni visive, dalle più elementari a quelle di tipo più complesso, che è molto simile a quella che può verificarsi in risposta a certe sostanze (ad esempio la mescalina), in seguito a privazione del sonno, o a privazione sensoriale. Si può confrontare la sequenza emicranica con quanto riporta Hebb sulle allucinazioni visive indotte da privazione sensoriale:
«Sembra che l’attività abbia un corso di sviluppo piuttosto regolare, dal semplice al complesso. Il primo sintomo è che il campo visivo, quando gli occhi sono chiusi, passa dall’oscurità al chiarore; viene poi descritta la comparsa di punti, linee, motivi geometrici ... Il passo successivo è la visione di qualcosa che somiglia ai motivi di una carta da parati ... Poi compaiono oggetti isolati, senza sfondo ... infine, scene integrate che presentano, di solito, distorsioni simili a quelle dei sogni» (Hebb, 1954).
Le configurazioni e il movimento dei fosfeni emicranici più semplici attraverso il campo visivo ricordano le allucinazioni cromatiche di forma astratta (luci lampeggianti, stelle, ruote, dischi, sfere roteanti, ecc.) che Penfield e Rasmussen consideravano evocate dalla stimolazione diretta della corteccia visiva esposta (area 17).
L’aspetto degli scotomi scintillanti sembra essere limitato al processo emicranico e nessuna procedura sperimentale ha consentito (ancora) di simularlo. Lashley (1941) ha ipotizzato che la caratteristica microstruttura di questi scotomi (minutamente angolata, e più grossolana nelle parti inferiori del campo visivo, come mostra la figura 12) sia in relazione con il disegno citoarchitettonico, o grana neuronica, della corteccia visiva primaria.
Lashley ha anche considerato la velocità di diffusione degli scotomi: secondo i suoi calcoli, un’onda di eccitamento si propaga sulla corteccia visiva (primaria) alla velocità di 3 mm al minuto, seguita da un’onda di totale inibizione. Egli ammette che «non si sa nulla dell’effettiva attività nervosa» alla base di questo processo, e perciò non si schiera, né con la teoria di un disturbo originato a livello locale, né con quella di un disturbo secondario in risposta a un’eccitazione proveniente da livelli infracorticali.
Non è chiaro perché la corteccia visiva debba essere più sensibile alla stimolazione delle aree corrispondenti della corteccia cutaneo-cinestetica (area 3) o della corteccia uditiva (area 41); e nemmeno è chiaro quale processo fondamentale sia riflesso nella frequenza di scintillazione degli scotomi o delle parestesie (da 6 a 12 al secondo). È probabile, tuttavia, che queste scintillazioni siano della stessa frequenza delle onde alfa e delle luci stroboscopiche che provocano il trascinamento fotico dell’elettroencefalogramma, la fotoepilessia e i fotoscotomi. È lecito sospettare che questa frequenza sia in rapporto con una velocità definita di elaborazione o scansione percettiva.
Dopo il verificarsi di semplici fosfeni e di scotomi scintillanti, possono seguire percezioni visive erronee e allucinazioni di ordine superiore: visione lillipuziana e brobdignagiana, varie forme di agnosia visiva, visione a mosaico, sequenze stereotipate (cinematiche) di immagini visive. È stato stabilito sperimentalmente (Penfield e Rasmussen, 1950) che la stimolazione dei campi secondari o periferici della corteccia visiva può generare allucinazioni visive organizzate, disposte in una sequenza spazio-temporale definita, e possiamo ipotizzare che tutte le allucinazioni visive più complesse insorgano a questo livello (o a un livello superiore). Riguardo ai fenomeni della visione a mosaico e alle dimensioni variabili delle tessere percepite (si veda la figura 14) è chiaro che si può ipotizzare una qualche forma di schematizzazione al di là dei modelli citoarchitettonici prefissati anatomicamente.
Konorski (1967) ha proposto e argomentato una teoria, fondata su unità percettive o gnosiche (strutture minime della percezione), che potrebbe avere attinenza con i fenomeni della visione a mosaico. Abbiamo detto che quest’ultima comincia come una granulosità appena percettibile, o un’apparenza minutamente cristallina; poi, attraverso stadi successivi in cui le dimensioni dei grani vanno crescendo, si arriva all’agnosia visiva. Si può supporre che questi sintomi rappresentino l’esperienza soggettiva di unità gnosiche via via più grandi che, normalmente invisibili, si impongono alla coscienza con l’aumentare delle dimensioni, presentandosi come unità poligonali sempre più ampie, fino a che la dimensione supera il loro contenuto d’informazione – e a questo punto il riconoscimento degli oggetti diviene difficile o impossibile, come in una fotografia con una grana troppo grossa.
Le allucinazioni sensoriali più complesse dell’aura emicranica assumono la forma di sinestesie e di altre interazioni sensoriali, di sequenze simili a sogni che coinvolgono immagini sensoriali di tutti i tipi, di afasie recettive ed espressive, di disordini generali del pensiero e del comportamento.
Si può pensare che questa gerarchia di allucinazioni osservata nelle aure emicraniche sia correlata all’attivazione successiva di campi corticali diversi. I campi corticali centrali (ad esempio l’area 17 nella corteccia visiva) sono «distinguibili dagli altri campi per la “grossolanità” della loro struttura neuronica, che è adattata alla ricezione e all’emissione di intensi flussi di eccitazione» (Lurija, 1966). Questi campi centrali, soprattutto quelli della corteccia visiva, sembrano essere più sensibili a un’eccitazione proiettata rostralmente, e la loro risposta è avvertita come semplici allucinazioni somatotopiche dei campi visivo o tattile (scotomi e parestesie); se l’eccitazione è forte, essa può diffondersi nei campi sensoriali secondari, dando qui origine a deficit (agnosia) e ad allucinazioni più complesse di un’unica modalità; nelle aure emicraniche più intense possono essere attivati anche i campi corticali terziari, che sono associati – secondo le parole di Lurija – con «le forme più complesse di integrazione dell’attività congiunta degli analizzatori visivi, uditivi e cinestetici»; la loro stimolazione dà origine alla disorganizzazione delle manifestazioni più generali dell’attività corticale, e cioè a stati confusionali complessi. La figura 18 mostra le aree della corteccia occupate dai campi primari, secondari e terziari, le loro sovrapposizioni e i loro confini reciproci.
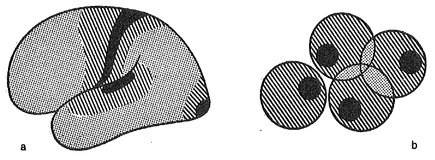
Fig. 18. Relazione tra aura emicranica e aree corticali. Si ritiene che le allucinazioni sensoriali elementari (visive, tattili, uditive) insorgano per eccitazione dei campi neuronici primari (in nero) della corteccia cerebrale, caratterizzati da un basso livello di soglia, e che le alterazioni più complesse della funzione percettiva e integrativa insorgano per il diffondersi di una forte eccitazione nei campi secondari (fondo rigato) e terziari (fondo punteggiato). (Da Poljakov, ridisegnato).
Sappiamo che nelle emicranie la gamma dei sintomi è vasta: si estende dai disturbi vegetativi elementari (con interessamento dei plessi autonomi periferici) attraverso disturbi dei meccanismi di risveglio centrale, fino a una varietà di disturbi corticali che interessano diversi ordini di campi neuronici. Questo è il repertorio dell’emicrania. Dobbiamo ora occuparci del problema delle sue variazioni e della variabilità entro questa gamma. Ad esempio, le emicranie comuni non mostrano alcun sintomo di interessamento corticale, mentre aure emicraniche isolate presentano principalmente sintomi di questo tipo; nelle emicranie classiche tale interessamento può manifestarsi a diversi livelli.
Dobbiamo indagare le basi fisiologiche delle permutazioni e delle trasformazioni che sono tanto caratteristiche dell’emicrania: i passaggi da un equivalente emicranico a un altro, dalle nevralgie emicraniche alle emicranie comuni, dalle emicranie alle epilessie, ai mancamenti, agli attacchi vagali, a tutte le altre reazioni associate che Gowers classificherebbe nella stessa zona di confine. Questi sono gli aspetti più difficili e affascinanti dell’emicrania; è chiaro che non possiamo neanche avvicinarci a essi se non adottiamo alcuni concetti radicalmente diversi sul funzionamento cerebrale. È impossibile spiegare i livelli variabili e le trasformazioni dell’emicrania in termini di meccanismi neurali fissi.
Già un secolo fa lo avevano compreso chiaramente sia Liveing, sia Jackson; entrambi arrivarono a risposte simili, sebbene quella di Jackson fosse elaborata in modo assai più cauto. Liveing asseriva che «... la concentrazione di tale tendenza [l’energia nervosa] in particolari distretti ... sarà il principale determinante del carattere della nevrosi in questione». Jackson fu costretto a rinunciare all’idea di centri anatomici unici e di funzioni anatomicamente localizzate; egli immaginò invece che il sistema nervoso fosse organizzato in modo gerarchico, e consistesse di diversi livelli nei quali ogni funzione fosse rappresentata:
«Io faccio l’ipotesi che il sistema nervoso sia un meccanismo sensomotorio, diretto dal basso verso l’alto; che ogni parte del sistema nervoso rappresenti sensazioni o movimenti, o entrambi ... La periferia è il vero livello inferiore; tuttavia parleremo di tre livelli di evoluzione centrale. 1) Il livello più basso è costituito dalle corna anteriori e posteriori del midollo spinale ... e dagli omologhi di tali parti, più rostralmente ... 2) Il livello intermedio è costituito dalla regione motoria di Ferrier, con i gangli del corpo striato, e ne comprende anche la regione sensitiva. Esso rappresenta tutte le parti del corpo in modo doppiamente indiretto. 3) Il livello più alto è costituito dai centri superiori motori (lobi prefrontali) e sensitivi (lobi occipitali)».
Egli assegnava quindi a ogni funzione un’organizzazione «verticale» complessa: la localizzazione di un sintomo non poteva in alcun modo essere identificata con la localizzazione della particolare funzione compromessa. Epilessie diverse dimostravano una dissoluzione gerarchica della funzione: ad esempio, un accesso di grand mal era riconosciuto come un attacco del livello superiore, e un accesso di laringospasmo (emicrania o epilessia laringea) come un attacco del livello inferiore. Jackson dedicò poco spazio a una discussione dell’emicrania, ma è chiaro che considerava solo gli attacchi classici quando scriveva:
«Credo che alcuni casi di emicrania siano in realtà epilessie (epilessie sensoriali) ... Penso che i sintomi sensoriali del parossismo siano dovuti a una “lesione scaricante” delle circonvoluzioni evolutesi dal talamo ottico, e cioè dei “centri sensitivi medi” ... Credo che il mal di testa e il vomito siano fenomeni postparossistici».
Le emicranie comuni, con i loro sintomi prevalentemente vegetativi, sarebbero considerate come forme di attacco del livello inferiore; le aure emicraniche, accompagnate da allucinazioni complesse e stati di assenza, come attacchi del livello superiore. Tutte le forme di emicrania, in termini jacksoniani, condividono la stessa organizzazione, espressa attraverso meccanismi omologhi a livelli diversi.
Nella sua analisi delle epilessie, Jackson si interessò soprattutto della rappresentazione gerarchica del movimento e delle funzioni motorie entro il sistema nervoso, mentre noi siamo interessati all’organizzazione verticale delle funzioni vegetative e sensoriali. Il fatto che anche queste siano rappresentate e rirappresentate a livelli progressivamente superiori nel nevrasse fornisce, per così dire, un’altra dimensione nella scelta della forma clinica degli attacchi. La sequenza o la sindrome emicranica può essere espressa al livello jacksoniano superiore (come un’aura complessa), al livello jacksoniano intermedio (come un’aura elementare, che interessa solo i campi sensitivi primari della corteccia), o al livello jacksoniano inferiore (come un’emicrania comune o un equivalente emicranico). Jackson parla anche di diffusione collaterale, come pure di diffusione verticale, nell’origine delle epilessie, là dove la prima espressione si riferisce all’interessamento di un’area contigua allo stesso livello funzionale. L’assenza di un «focus scaricante» anatomicamente definito significa che questa diffusione laterale non ha restrizioni: ad esempio, un’aura che scaturisce dall’attivazione dei campi sensitivi primari può interessare o «scegliere», indiscriminatamente, il campo tattile o quello visivo, mentre un’emicrania comune del livello inferiore può interessare una qualunque porzione dei campi vegetativi al livello del tronco cerebrale, dando luogo, con la stessa facilità, ora a un attacco di mal di testa, ora a un attacco addominale, ora a un attacco precordiale.
Sono gli studiosi postpavloviani di riflessologia che soprattutto si sono dedicati a verificare ed estendere le idee di Jackson sulla funzione e la localizzazione nel sistema nervoso; ciò ha comportato una radicale ridefinizione e il ripensamento di molte dottrine fondamentali (si veda Lurija, 1966). Ad esempio, secondo questa scuola una funzione è, in realtà, un sistema funzionale orientato verso l’esecuzione di un particolare compito biologico. Tale sistema funzionale si basa – e questo è l’aspetto più significativo – su una «costellazione» dinamica di connessioni, situate a diversi livelli nel sistema nervoso, le quali possono essere liberamente sostituite o scambiate l’una con l’altra, fermo restando il loro compito. Così, con le parole di Lurija, «un tale sistema di componenti in legame funzionale ha una struttura sistematica e non concreta, nella quale l’anello iniziale e finale del sistema (il compito e l’effetto) rimangono costanti e immodificati, mentre gli anelli intermedi (i mezzi per eseguire il compito) possono essere modificati entro ampi limiti».
Queste considerazioni, discusse da Lurija a proposito del movimento e dei compiti motori, si presentano come indispensabili per comprendere l’emicrania e le sue trasformazioni come compiti vegetativi e psicosomatici. Il compito può essere foggiato dalla necessità di una scarica neuronica (in un’emicrania periodica o parossistica), oppure per necessità fisiche o emotive, come negli attacchi circostanziali o abituali; l’anello finale, l’effetto dell’emicrania, è rappresentato dal ripristino di un equilibrio fisiologico (o emotivo). Tuttavia, il compito adattativo ha una struttura sistematica e non concreta: ciò significa che i meccanismi effettivamente impiegati possono essere molti, diversi e non costanti. Ci possono essere tanti modi di costruire un’emicrania quanti di cucinare un’omelette. Se viene eliminato un particolare anello intermedio, un ingranaggio, l’intero sistema può riorganizzarsi in modo da ripristinare il compito disturbato.
Così, come accade per i compiti motori o percettivi, i meccanismi particolari sono subordinati alla strategia complessiva. Questo principio ha grande importanza pratica e terapeutica, oltre che un interesse teorico. Esso significa, ad esempio, che se nell’economia fisiologica o psicologica di un individuo un’emicrania è necessaria, gli attacchi continueranno a verificarsi e ad essere elaborati, quale che sia il particolare meccanismo eliminato. Si recida pure un’arteria temporale, o una formazione terminale, e un’altra verrà messa in azione; si cerchi di bloccare gli attacchi somministrando, ad esempio, un inibitore della serotonina, ed è probabile che gli attacchi si ripresentino utilizzando un diverso meccanismo intermedio.
Sistemi funzionali come questi, sostiene Lurija, dalla composizione complessa, plastici nella variabilità dei loro elementi, e in possesso della capacità di autoregolarsi dinamicamente, sembrano essere la norma, nell’attività umana.
Ricapitolazione
La reazione emicranica è caratterizzata, ai livelli funzionali inferiori, da un tono parasimpatico o trofotropico protratto, preceduto e seguito da stati fisiologicamente opposti. Ai livelli funzionali superiori, essa è caratterizzata dall’attivazione (e dalla successiva inibizione) di innumerevoli campi corticali, dalle aree sensoriali primarie alle aree integrative più complesse. L’emicrania è considerata una forma di parossismo centroencefalico in lenta evoluzione: nel caso dell’aura l’evoluzione è da 20 a 200 volte più lenta e nel caso dell’emicrania comune alcune migliaia di volte più lenta delle rispettive controparti epilettiche. Bisogna anche considerare l’emicrania come un compito adattativo complesso eseguito da un sistema funzionale complesso, nel quale i mezzi di esecuzione (che sono estremamente variabili) sono subordinati ai suoi fini.
L’emicrania sfuma in una diffusa regione di confine nella quale troviamo reazioni parossistiche e adattative associate. Se ne può dare una rappresentazione schematica in un diagramma che riporta sull’asse delle ordinate il livello funzionale (jacksoniano) e su quello delle ascisse la durata di tali reazioni (figura 19). Dovrebbe esserci anche un terzo asse, indicante le diverse sindromi dell’emicrania e delle reazioni associate per una data scala temporale e per un dato livello funzionale.
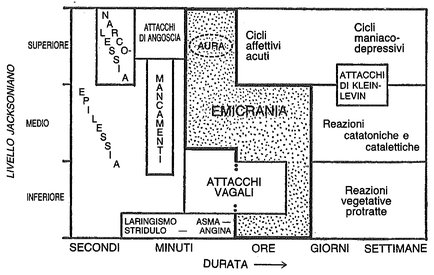
Fig. 19. Relazioni tra emicrania e alcuni disturbi associati, rappresentati in termini di durata temporale e di livello neurale (jacksoniano). Entrambi questi parametri possono variare opportunamente, e ciò spiega le metamorfosi cui l’emicrania è suscettibile. Tutti i disturbi qui rappresentati sono distinti e individuati, ma nelle zone di separazione sconfinano gli uni negli altri.
Per gli scopi pratici (diagnostici e terapeutici), possiamo identificare la maggior parte delle emicranie come emicranie e nient’altro; teoricamente, invece, possiamo aspettarci forme ibride e attacchi di transizione di ogni sorta, là dove l’emicrania confluisce nelle epilessie, nei mancamenti, negli attacchi vagali, e in altre crisi vegetative ed emotive.42 Di tanto in tanto accade di imbattersi in questi attacchi ibridi di transizione, che vanificano (e in effetti, rendono priva di significato) la fatica della diagnosi differenziale.
Si deve sospettare che i confini fisiologici non siano più precisi, in questo contesto, di quelli clinici, e che pertanto non sia altro che una chimera la ricerca di un «processo emicranico» unico e patognomonico nel sistema nervoso. Non possiamo immaginare che la ricerca sia in grado di mettere a nudo un’anormalità semplice e netta come i complessi onda-picco nel caso di certe epilessie. Dobbiamo invece prevedere un ampio spettro di attività nervose espresse a differenti livelli funzionali e su differenti scale temporali.